


快速眼动 (REM) 睡眠
是一种在睡眠状态下,脑电波表现为低幅快波,因为与觉醒时的脑电波相似,也称为快波睡眠。
并且,由于脑电波显示皮层活跃,实际处于睡眠状态,因此又称为异相睡眠。
快速眼动睡眠期间,人体内部的许多生理功能会发生明显变化, 如心率减慢、血压下降、新陈代谢缓慢、呼吸次数减少。其特点以机体感觉减退、肌肉放松、交感神经活跃性降低和下丘脑体温调节功能减退等表现为主。
在这个阶段常有梦境形成,74%-95%的人群在此睡眠阶段会做梦,但*7%左右人群能够清晰回忆梦中的情景。
迄今为止,REM睡眠对机体的生理和心理机能尚有很多内容等待我们探索。但研究者根据各种实验结果,REM已被证明参与复杂概念记忆以及情感记忆、视觉和运动程序的调节回忆,它对于记忆巩固发挥着重要的作用。
在REM 期间,基因表达的上调可能促进海马体的长期增强,以及皮层突触可塑性的重组。因此,假设有一种操纵快速眼动睡眠的方法能够被开发出来,这将为更好地理解这个睡眠阶段在记忆和认知中的各种作用提供一个宝贵的工具。
早期研究表明,快速眼动时的听觉刺激可能会调节快速眼动时间并降低快速眼动密度。在这基础上,该篇文献研究了与此类刺激相关的认知效应和脑电图反应。
20 世纪 80 年代和 90 年代的一系列研究提供了初步证据,表明通过感觉刺激改变 REM 的电生理特征是可能的。
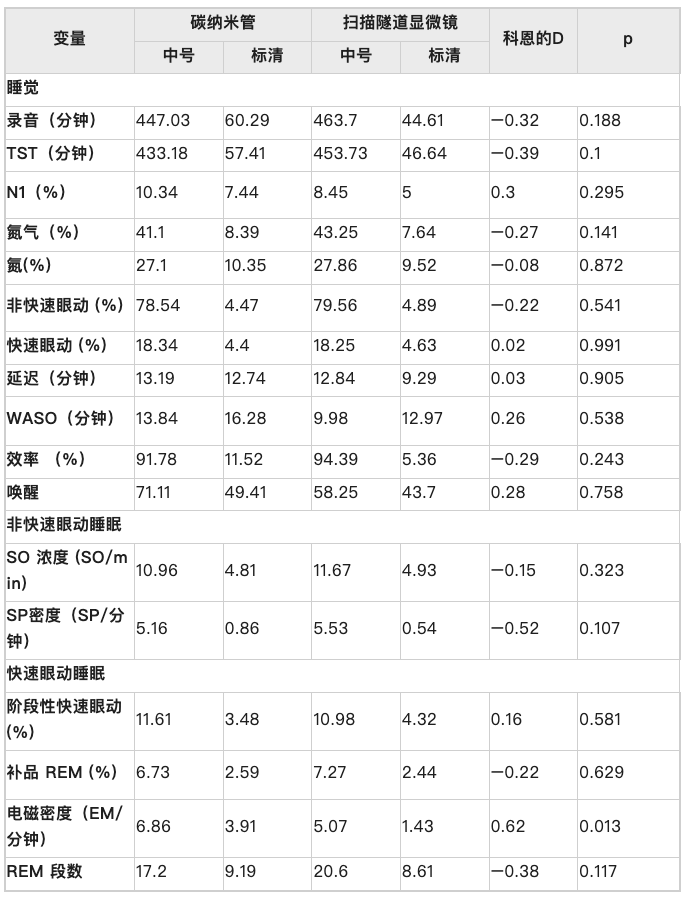
这些早期研究表明,由[1,2,3]REM 眼动 (EM) 触发的短暂听觉刺激可以增加人类 REM 睡眠的长度,同时也降低 EM 的密度。重要的是,许多研究还设法通过未锁定 EM 的听觉刺激来操纵 REM,例如 refs[4,5,6]。在大多数情况下,这是成功的,并且增加了 REM 持续时间,同时减少了 EM。[1] Mouze-Amady, M.、Sockeel, P. 和 Leconte, P. REM 偶然听觉刺激对人类 REM 睡眠行为的修改。生理学。行为。37、543-548(1986)。
[2] Sockeel, P.、Mouze-Amady, M. 和 Leconte, P. 人类快速眼动睡眠期间听觉生物反馈回路引起的脑电图不对称性的改变。国际。J.心理生理学。
[3] Guerrien,A.,Dujardin,K.,Mandal,O.,Sockeel,P. 和 Leconte,P。人类学习后快速眼动睡眠期间听觉刺激增强记忆。生理学。行为。45、947-950(1989)。
[4] Salin-Pascual, RJ、Jimenez-Anguiano, A.、Duran-Vazquez, A.、Nancy, HM 和 Drucker-Colin, R. REM 睡眠剥夺后恢复期间听觉刺激的管理。睡眠 17 , 231–235 (1994)。
[5] Vazquez, J.、Merchant-Nancy, H.、Garcia, F. 和 Drucker-Colin, R. 感觉刺激对快速眼动睡眠持续时间的影响。睡眠21 , 138–142 (1998)。
[6] Arankowsky-Sandoval,G.,Stone,WS 和 Gold,PE 通过听觉刺激增强年轻和年老大鼠的快速眼动睡眠。脑研究。 589、353-357(1992)。
然而,[3]对 EM 关联方法和非 EM 关联方法的直接比较表明,在促进莫尔斯电码比较任务的记忆巩固方面,前者明显更有效。这是**一项研究这种刺激对记忆巩固影响的研究。在当前的研究的基础上,该篇文献探索了电磁触发的听觉刺激如何主动改变快速眼动电生理学,以及这种刺激如何影响非听觉快速眼动依赖性记忆任务的巩固,这些任务的重放不能由点击刺激触发。如果这些假设成功,研究可以预测听觉刺激将导致脑电图活动的变化,从而预测认知表现。
(i) 注意力任务(n=20):精神运动警惕性测试(PVT)
(ii) 视觉任务(n=18):纹理辨别测试(VDT)
(iii) 程序性任务(n=20):镜像追踪测试(MTT)
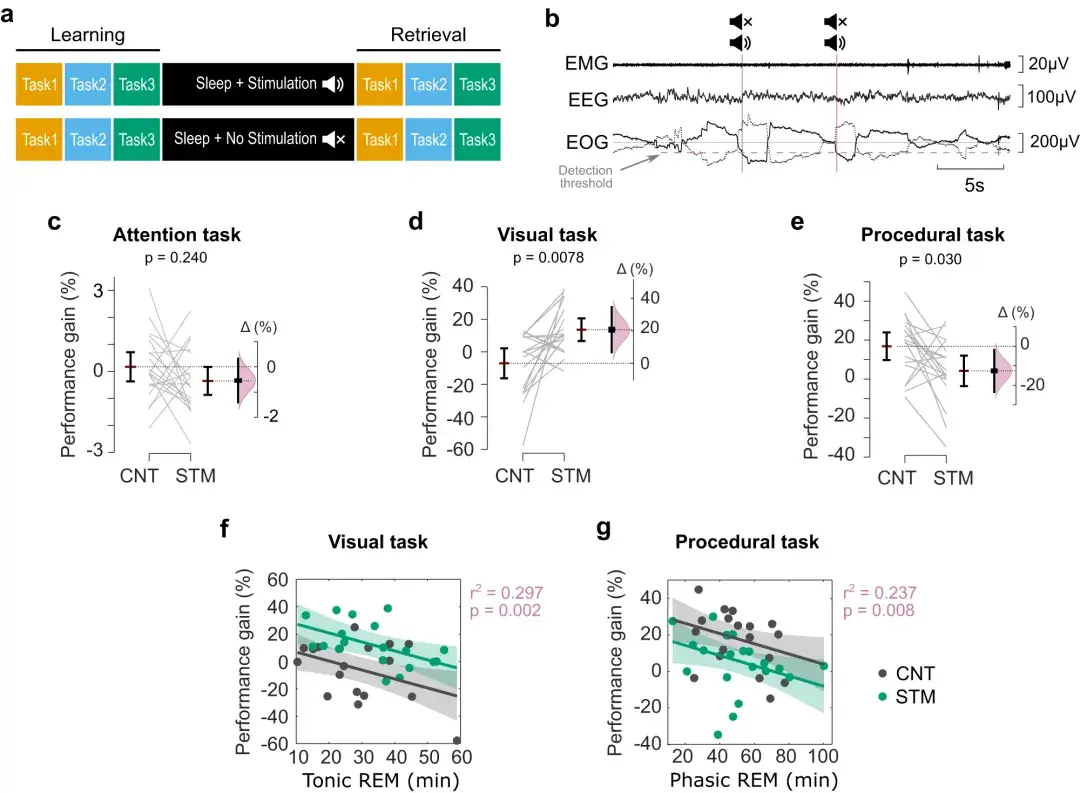
实验中,参与者睡眠约7小时,并且在刺激程序(STM)的REM期间使用锁定在眼动(EM)上的咔嗒声,或者在控制程序(CNT)中静音声音。
睡眠后,参与者接受了同样的认知任务测试。睡眠的顺序(CNT, STM)和任务的顺序在参与者之间是随机的,但在参与者的睡眠前和睡眠后测试中,任务的顺序是相同的。
持续监测一条EOG通道,在稳定REM睡眠时**EM检测。手动设置一个振幅阈值(50-100 μ V),当EOG振幅越过这个水平时,会发出咔嗒声,然后刺激暂停2.5 s。在 CNT 条件下,咔嗒声会被静音。
视觉 (VDT) ( d ) 和程序 (MTT) ( e ) 任务中夜间性能增益的变化以及 STM 和 CNT 条件的效应大小。
多变量线性模型表明,强直性 REM 花费的时间可以预测视觉任务表现的夜间变化(增益百分比 = 13.5 + 20.5 C–0.66tR)( f)。然而,阶段性 REM 花费的时间可以预测程序性任务的表现增益(增益百分比 = 31.9.5–12.0 C–0.28pR) ( g )。
误差线表示平均值 ±95% CI,Δ 通过平均值差异显示效应大小。效应大小的分布曲线表示给定观测数据的 Δ 的重采样分布。
对于 f 和 g 中的多元线性回归:C = 条件 (CNT/STM),tR = 强直 REM 和 pR = 阶段性 REM,阴影对应于响应的 95% CI。
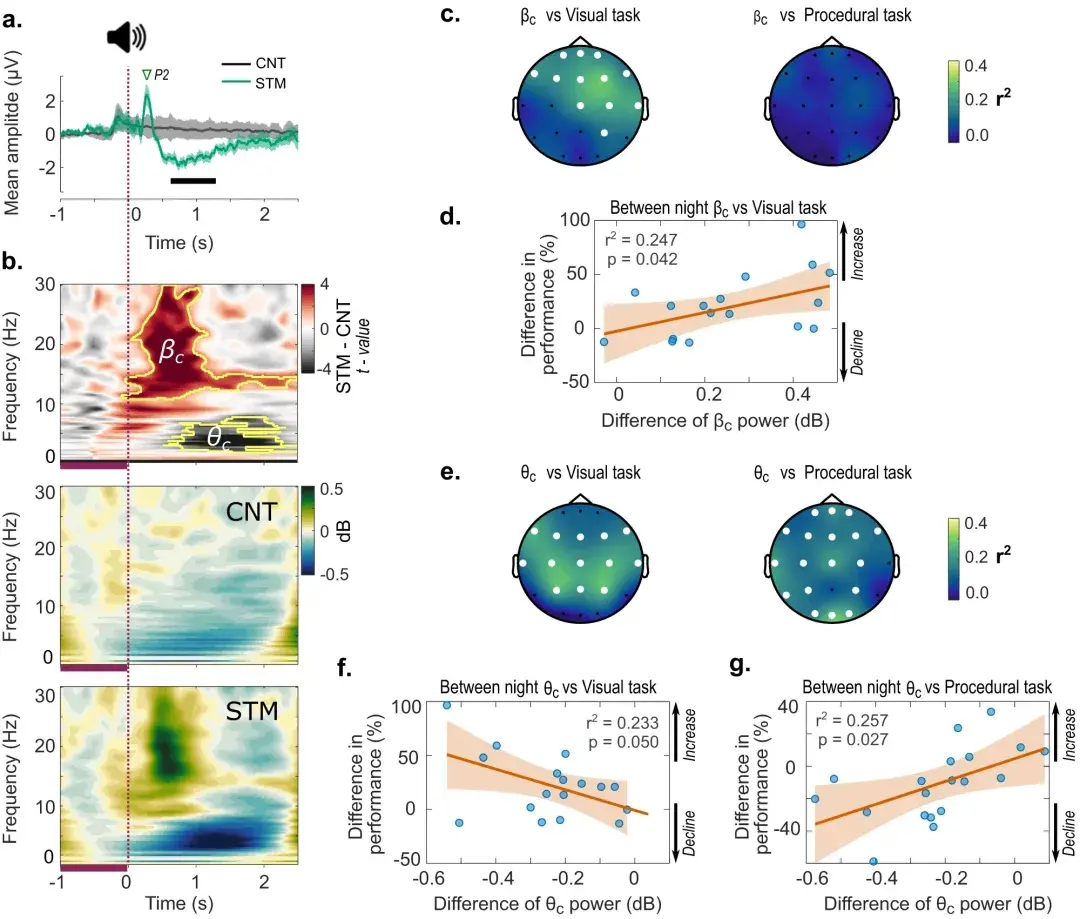
接下来进行电生理分析,研究使用所有通道的头皮 ERP 和 ERSP 评估了头皮 EEG 对听觉刺激的反应。
a 事件相关电位 (ERP) 在所有通道中平均,以响应 REM 期间锁定到 EM 的听觉刺激。粗黑条表示条件之间存在显着差异。阴影表示平均值±95% CI。
b 所有通道响应听觉点击的平均事件相关频谱扰动 (ERSP)。
对条件之间光谱差异的分析表明,β ( βc ) 和 theta ( θc ) 频率存在两个主要聚类(p < 0.05,聚类校正)。时频图下方 -1 和 0 s 之间的深紫色条表示频率功率 (dB) 变化的基线周期。
每个电极的βc ( c ) 和θc ( e )内的功率变化符合线性回归,预测视觉 (VDT) 和程序 (MTT) 任务性能的夜间改善。白色通道表示 FDR 校正后显着的线性拟合 ( q < 0.05),表明行为表现和集群权力之间存在线性关系。因此,夜间相对功率差异(对于重要通道)(CNT-vs.-STM)与条件之间视觉任务的夜间性能增益相关( d )。
同样,条件间(CNT-vs-STM)相对θc功率(对于重要通道)的差异与视觉任务 ( f )夜间表现的下降略有相关,而θc功率的相同差异与性能的提高相关对于程序任务 ( g )。CNT 与 STM 的性能差异为正值表示 STM 夜晚的性能提高,而负值表示 STM 夜晚的性能下降。同样,CNT 与 STM 的功率差异大于 0 表明 STM 中的功率较大,而功率的变化小于 0 表明 CNT 中的功率较大。阴影对应于 ( d、f 和 g )中回归图响应的 95% CI 。
结论
结果表明,快速眼动睡眠中的这种听觉刺激会减少快速眼动睡眠的特征,并提高视觉任务的表现,但不利于镜像追踪任务。
研究还观察到刺激后β带活性增加和θ带活性减少。并且,这些光谱变化与行为表现的变化有关。
所以,声音刺激可以调节快速眼动睡眠,并表明不同的记忆过程支撑着其对认知表现的不同影响。
CONTACT 联系我们
联系人:樊女士
电话:18900616086 京显
邮箱:18900616086@163.com changxian-el@hotmail.com
地址:北京市海淀区中关村南大街5号二683号楼
联系方式▼ 更多咨询关注小程序▼


 产品展示
产品展示


 http://bitbrain.cn/Zz_www.eastsummit.net/index.html
http://bitbrain.cn/Zz_www.eastsummit.net/index.html 
 销售一部
销售一部